



L'increïble (i amagat) món dels insectes I
-
- Home
-
- 14 of 24

És indubtable que hi ha centenars de milions d'insectes que ens envolten i és espectacular analitzar la seva capacitat per adaptar-se i poder-los trobar en qualsevol hàbitat al nostre planeta [3-4]. Això és gràcies a la seva increïble capacitat per modificar les seves propietats a partir del control de la seva micro i nanoestructura que genera grans canvis en moltes propietats com l'adhesió, la locomoció o les seves propietats físiques i químiques (òptica, fotònica, hidrofobicitat, sensors i actuadors mecànics, acústics, òptics, tèrmics i químics).
La teoria de les espècies: els insectes com a paradigma
Una multitud de pressions selectives acompanyen la diversitat dels insectes, i els ha portat a desenvolupar solucions complexes i efectives per facilitar la seva supervivència i adaptabilitat. Els insectes exhibeixen enlluernadores propietats òptiques com a resultat de cristalls fotònics naturals, patrons jeràrquics precisos que abasten escales de longitud des de nanòmetres fins a mil·límetres, i mecanismes de defensa formidables. Aquesta complexa evolució ha bioinspirat els humans per desenvolupar sistemes o aplicacions molt interessants que, per exemple, inclouen recobriments antireflectors, pantalles òptiques i algorismes de computació.

Els insectes mereixen una atenció particular des d'una perspectiva bioinspiracional atesa la seva sorprenent abundància i diversitat, perquè representen la meitat de les espècies del planeta; possiblement representen l'èxit més salvatge de la selecció natural. Podem trobar més d'un milió d'espècies descrites en la classe Insecta, amb estimacions que van des de tres milions fins a desenes de milions. Només els escarabats ronden les 240.000 espècies conegudes (en comparació, hi ha només al voltant de 6.000 espècies de mamífers conegudes) [5], i en un sol arbre al Perú es van trobar 43 espècies diferents de formigues [6]. Els insectes són a tot arreu i s'han adaptat a gairebé tots els ambients de la Terra, encara que només un grapat d'espècies viuen en els oceans o a les regions polars. En conseqüència, els insectes ocupen molts rols en la cadena alimentària mundial, mengen plantes vives i mortes, fongs, altres insectes i vertebrats, i serveixen com una font d'aliment comú per als animals superiors, inclòs l'Homo sapiens [3, 7].

Els insectes: productes nanotecnològics
La capacitat dels insectes per adaptar-se a diversos entorns està vinculada a dos sistemes evolutivament optimitzats: un exoesquelet derivat de la cutícula amb micro i nanoestructures funcionals associades, i els complexos glandulars que secreten substàncies químicament diverses. La majoria de les estructures funcionals depenen profundament de l'organització jeràrquica, amb ordenament estructural en escales de longitud que van de nanòmetres a mil·límetres. Les adaptacions estructurals dels insectes funcionen per satisfer un ampli conjunt de necessitats dels insectes, incloent-hi la detecció i el control ambiental, la protecció, la comunicació i la locomoció.
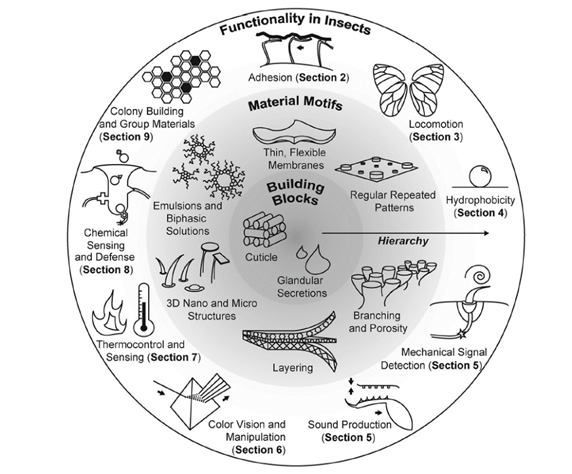
Figura 1. Anàlisi de l'efecte que tenen les micro i nanoestructures en les propietats dels insectes
Adhesió
Al començament del segle XVII, poc després de la invenció del microscopi, els investigadors van poder estudiar els detalls dels mecanismes adhesius per primera vegada. Ràpidament, es va iniciar una animada discussió sobre com els insectes podien adherir-se i caminar sobre molts tipus de superfícies. Al llarg dels últims dos segles, s'han proposat moltes hipòtesis sobre l'adhesió d'insectes, que van des de les “esponges farcides de glútens”, a l'entrellaçament de pèls, passant per les ventoses i fins i tot per les secrecions adhesives (Fig. 2a i 2b) [8-10].

Figura 2 a) Un dels primers dibuixos dels peus adhesius d'una mosca publicat per Robert Hooke en 1665. "El peu d'una mosca [...] és d'una invenció molt admirable i curiosa" (Figura adaptada de [8], domini públic). (b) En viatjar a la terra de Brobdingnag, el capità Lemuel Gulliver és atacat per vespes gegants, "Rastrejant aquesta matèria viscosa que, segons ens diuen els nostres naturalistes, permet a aquestes criatures caminar amb els peus cap amunt sobre un sostre" (Figura adaptada de [9], domini públic)
Podem dividir en dos els sistemes adhesius dels insectes:
a) Estructures adhesives físiques que s'entrellacen mecànicament o generen una força atractiva a través de les interaccions de Van der Waals, sent sistemes similars als que presenta la flor de lotus, és a dir, la presència de micro i nanoestructures que aporten superhidrofobicitat i autoneteja a les fulles de nelumbe (Fig. 3a).
b) Les secrecions d'adhesius químics que actuen a través d'enllaços moleculars, forces capil·lars i forces viscoses.
Sistemes que els permeten, per exemple, adherir-se ràpidament a les superfícies (i sovint desprendre-se'n) o la capacitat d'autonetejar-se, i així eliminar contaminants. Les estructures fibril·lars o ramificades són les més habituals en els sistemes adhesius que es troben en els insectes. Així com els sistemes basats en interaccions com l'enllaç d'hidrogen, les forces viscoses i les forces de Van de Waals (Fig. 3b). És important remarcar que gràcies a la comprensió d’aquests sistemes d’adhesió, ja tenim productes en el mercat bioinspirats que aporten interessants solucions per millorar l’adhesió o per crear l’efecte d’autoneteja [11].

Figura 3. a) Imatges de l’efecte lotus, la fulla de lotus, i imatges de SEM i representació de les micro i nanoestructures [12]
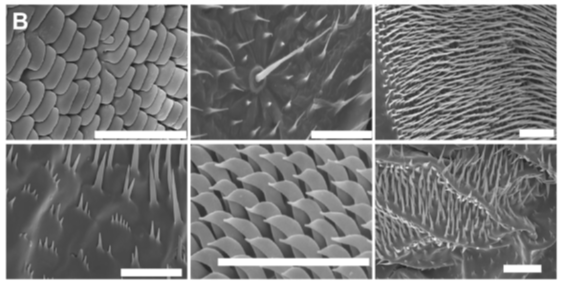
Figura 3 b) Estructura lamel·lar de la cutícula de l'insecte, que forma la majoria de l'exoesquelet d'un insecte. Tres regions diferents de capes no vivents descansen sobre una capa epidèrmica viva que alberga diverses cèl·lules relacionades amb la microestructura i productores de cutícules. La capa més externa d'epicutícula representa la primera línia de defensa entre l'insecte i el seu entorn extern. Sovint conté secrecions de lípids, ceres o altres recobriments per manipular la humectabilitat, potser les més intrigants de les quals són nanopartícules intricadament estructurades, conegudes com a brocosomes, que es troben en les llagostes de la família Cicadellidae. Sota aquesta capa es troba l'exocutícula, que experimenta una reticulació extensa i té una rigidesa relativament alta. En molts insectes, s'endureix ràpidament per actuar com un embolcall protector després de la muda per al desenvolupament prolongat d'un endocúcul més suau i hidratat a sota. Les capes exo i endocuticulars formen col·lectivament el que es coneix com la procutícula, un material compost de proteïnes, polifenols, aigua i lípids. Juntament amb els arranjaments cristal·lins de la important i abundant polisacàrid lineal, la quitina. A) Adaptat de [13] Copyright 1982, Springer. B) Barres d'escala: 20 μm. Adaptat de [14] Copyright 2016, Springer.
La importància de les ales dels insectes
Prop de dotze mil espècies de vertebrats i més d'un milió d'espècies d'insectes han desenvolupat ales per desplaçar-se. En totes les espècies d'insectes voladors, les ales proporcionen tres funcionalitats clau:
- Actuen com a palanques, transmetent la força dels músculs en la base de l'ala a l'aire circumdant.
- Són perfils aerodinàmics que dirigeixen l'aire a través dels cops de l'ala per generar sustentació.
- Actuen com a bigues en volada, que es deformen sota una varietat de forces.
S'estima que l'ala d'un insecte té una resistència (mòdul de Young) de 2 i 5 GPa, comparable a la del niló, encara que aquest valor pot variar segons la localització de l'ala, les espècies d'insectes i l'estructura general de les ales. Les ales estan fetes de cutícula, que al seu torn està formada per tubs buits ramificats de diferents diàmetres i gruixos de paret, amb seccions transversals el·líptiques, circulars o en forma de campana per impartir una rigidesa de flexió dependent de l'axial. En general, les venes tenen un diàmetre major i parets més gruixudes prop de la base de l'ala on les tensions són més grans, després s'estreny cap a les puntes de les ales per reduir les forces d'inèrcia. Les venes tenen un paper important en la variació de la flexibilitat entre les vores anterior i posterior de les ales (Figura 4). Com a mètode per promoure més deformacions dins d'aquesta estructura venosa relativament rígida, la cutícula de certes ales d'insecte conté segments lineals flexibles, que actuen com a línies de plec [15]. Aquestes bandes es distribueixen independentment de les venes de suport; aquelles que es mouen radialment (de base a punta) mitjancen la flexió i la torsió, mentre que altres orientades transversalment (de vora a vora de sortida) actuen com a frontisses d'una via per ajudar les ales a doblegar-se i restablir-se després de completar una carrera descendent [15]. Altres línies flexibles romanen rígides durant els moviments de vol, però es deformen de manera reversible quan les ales entren en contacte amb els obstacles per evitar danys estructurals. Les membranes de les ales no només serveixen com un element deformable, sinó que també poden contribuir a la rigidesa general de la flexió. El seu gruix varia aproximadament quatre ordres de magnitud entre diferents espècies d'insectes, des de menys de 500 nm en les delicades ales de les mosques de la fruita (Drosophila sp.) fins a 1 mm en les robustes ales davanteres dels escarabats (ordre Coleoptera).
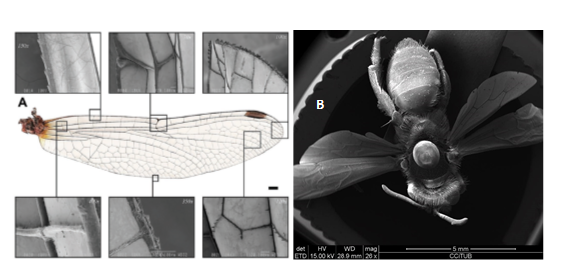
Figura 4. Les ales de libèl·lula estan avantatjosament venades i engrossides per proporcionar integritat estructural i variació espacial en la flexibilitat. A) Fotografia de la versió anterior de la libèl·lula vagabunda errant, Sympetrum vulgatum, amb imatges SEM associades a les diverses característiques estructurals de l'ala. Reproduït sota els termes de la llicència CC-BY [16]. Copyright 2010, Jongerius i Lentink i B) Imatges de SEM realitzades als Centres Científics i Tecnològics de la UB (CCITUB) d’una abella de l’espècie Osmia cedida pel Museu de Ciències Naturals de Barcelona.
Estructures associades a l’aigua
Les adaptacions que també es relacionen amb l'aigua es presenten en entorns que varien àmpliament en termes d'humitat, accés a la humitat i predomini de les interfícies, la qual cosa porta a una sèrie de pressions evolutives amb solucions diferents. Algunes d'aquestes solucions són externes, i les libèl·lules, cavallets del diable i els cigarrets tenen estructures cuticulars en les seves ales que protegeixen contra la interferència de les gotes de pluja i la brutícia. Unes altres són internes, per evitar la deshidratació.
Malgrat les seves diferències funcionals i espacials, la majoria de les adaptacions específiques de l'aigua en insectes comparteixen mecanismes d'acció comuns: modifiquen la humectabilitat a través d'una interfície diferent aire-aigua o mantenen pressions osmòtiques o hidrostàtiques específiques. Un cas curiós és l’escarabat de l'espècie Hoplia coerulea (figura 5), que pertany a la subfamilia Melolonthinae. Aquesta espècie té la capacitat de canviar de color i aprofitar-lo de diferent manera en funció de la captació d'aigua que fa en l'àmbit estructural [17]. Les escates a la cutícula de l'escarabat Hoplia coerulea poden absorbir aigua, amb la conseqüència que aquestes escates, que s'han demostrat que són responsables de la coloració blava brillant del coleòpter, de manera reversible es poden transformar en un color verd maragda amb un contingut d'aigua creixent. Les escates contenen una interessant estructura fotònica periòdica, composta d'un sòlid i una capa porosa composta de varetes rectangulars paral·leles. L’índex de refracció mitjà de tota l’estructura i el seu període vertical explica raonablement bé la coloració blava de l’insecte. El contacte de l’aigua a l’estructura de l’escarabat va omplint-la i modificant-ne el color. El mecanisme natural mostra la manera de produir un material higrocròmic molt eficient, és a dir, un mitjà que canvia significativament el color quan es modifica el contingut d'aigua.
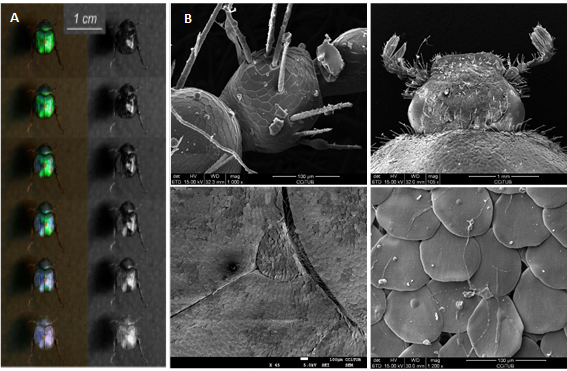
Figura 5 a) Es mostra en dues formes diferents una sèrie d’imatges fetes cada tres minuts durant el procés d’assecament, d'uns 15 minuts de durada. La sèrie esquerra no està processada, mentre que a la sèrie correcta (dreta), les imatges s'han processat de manera que no apareix la coloració de to verd fosc i la coloració blava sembla més lleugera. Adaptat de [17] b) Imatges de SEM a diferents augments realitzades als CCITUB d’escarabat de l'espècie Hoplia coerulea cedida pel Museu de Ciències Naturals de Barcelona.
Superfícies hidrofíliques i hidrofòbiques
La fulla de la flor de lotus és tot un paradigma dins de la naturalesa com a superfície superhidrofòbica i autonetejant [18-21]. Aquestes propietats són degudes a tota una sèrie de patrons en superfície jeràrquics micro i nanoestructurats (exemple a la fig. 6). Els insectes presenten característiques similars. Les superfícies tant d'algunes ales d'insectes com de les fulles de lotus estan esquitxades de micropilars cònics amb diàmetres en la base de 4 a 10 μm i altures d'entre 6 i 10 μm, separades entre 15 i 30 μm.
Aquestes micro i nanoestructures (i) eviten l'acumulació d'aigua (i, per tant, de pes) a través de la humectació, (ii) exhibeixen una baixa adherència a partícules estranyes, (iii) promouen la laminació de gotes per encapsular i eliminar qualsevol contaminant que aconsegueixi adherir-se a la superfície, (iv) estimulen coalescència de gotetes, que ajuda a protegir contra l'acumulació d'aigua de boires fines, i (v) dificulten el creixement bacterià.
En termes generals, aquests dissenys hidròfobs es poden generalitzar en almenys quatre grups: a) micro o nanoestructures simples (p. ex., amb forma de pilar o cúpula), b) micro o nanoestructures complexes (formes variades), c) escales (generalment de 2 a 3 μm en una dimensió), pèls o bolets molt més llargs (generalment més de 5 µm de llarg) que els seus diàmetres, i d) organitzacions jeràrquiques que inclouen qualsevol combinació d'aquests elements.
Figura 6 a) L'espècie col·lèmbols (Collembola) ha desenvolupat una varietat d'estructures superficials periòdiques i jeràrquiques amb propietats hidròfobes. A-E) Imatge d'Entomobrya intermèdia i imatges SEM que mostren motius hexagonals i triangulars en P. flavescens. B-F) Imatge de Vertagopus arboreus i imatges SEM que mostren motius quadrats i pentagonals irregulars en I. viridus. C-G) Imatge de Kalaphorura burmeisteri, i imatges SEM que mostren estructures granulars secundàries i motius hexagonals en S. quadrispina. D-H) Imatge de sp. ornata, i imatges SEM que mostren estructures granulars secundàries i patrons el·líptics variables en A. pygmaeus, barres d'escala: A – G = 2 μm, A –G = 500 nm. Reproduït amb permís [22] Copyright 2012, Springer.
Independentment del seu motiu de disseny, les estructures que indueixen a la hidrofòbia en els insectes generalment busquen maximitzar l'àrea d'interfície aire-aigua i minimitzar l'àrea de contacte aigua-sòlid. Aquest concepte d’hidrofobicitat va ser presentat per primera vegada per Cassie-Baxter en 1944 [23] per explicar la naturalesa hidrofòbica de les plomes d'ànec i com podrien servir de bioinspiració per a la roba repel·lent a l'aigua. Aquesta teoria descriu les micro i nanoestructures jeràrquiques com a superfícies corbes uniformes amb una composició heterogènia (p. ex., d'aire i sòlid), i postula que l'aire que omple l'espai entre aquestes estructures està essencialment atrapat i es comporta com un sòlid no humectant.
Un altre punt a tenir molt en compte per avaluar la importància d'aquestes estructures està relacionat amb la visió dels insectes. L'entelament per humitat es forma en una superfície quan les gotes de més de 190 nm de diàmetre (més de la meitat de la longitud d'ona més curta de la llum visible) es condensen i acumulen [24]. Aquest fenomen representa un desafiament per a la visió dels insectes perquè als insectes els manquen parpelles i, per tant, no tenen forma d'eliminar externament la humitat que bloqueja la visió o uns altres contaminants.
Els mosquits, família Culicidae, tenen una excel·lent visió que pot funcionar en ambients poc il·luminats i humits [25]. Per mantenir una visió constant i evitar l'entelament, la superfície de cada mosquit està recobert densament amb una espècie de tetines de grandària nanomètrica. Aquestes tetines tenen un diàmetre del voltant de 100 nm i estan espaiats aproximadament a 50 nm en una matriu hexagonal no empaquetada; també proporcionen un índex de refracció determinat per millorar-ne la transparència. Aquestes nanoestructures contenen espais d'aire que impedeixen que l'aigua entri en contacte amb una gran part de la superfície, tal com comentàvem en el model d'humectació de Cassie-Baxter [23,26-28].
Continuarà...
BIBLIOGRAFIA
[1] T. B. H. Schroeder; J. Houghtaling; B. D. Wilts; M. Maye. Adv. Mater. 2018, 30, 1705322
[3] J. M. Benyus. Biomimicry: Innovation Inspired by Nature. Perennial Nova York, 2002.
[4] Species 2000 & ITIS Catalogue of Life, 2016 Annual Checklist, http://www.catalogueoflife.org/annual-checklist/2016/browse/tree?068aebbdc768ab6cfa7d611d9f973e2d (consulta: setembre 2017).
[5] E. L. Mockford, Ann. Entomol. Soc. Am. 1997, 90, 115.
[6] J. Huber; J. Noyes; J. Hymenopt. Res. 2013, 32, 17.
[7] A. Jankielsohn. "The Importance of Insects in Agricultura Ecosystems". Advances in Entomology 06(02):62-73
[8] R. Hooke. Micrographia. Oxford Press, London (1665). Project Gutenberg, Public Domain
[9] J. Swift. Travels into Several Remote Nations of the World, in Four Parts. By Lemuel Gulliver, First a Surgeon, and then a Captain of Several Ships. Benjamin Motte, London (1726). Project Gutenberg, Public Domain
[10] J. Blackwall. Letter addressed to the secretary. Extracts from the Minute-Book of the Linnean Society 21:767 (1833)
[11] Nanotechnology solutions for self-cleaning, dirt and water-repellent coatings. https://bit.ly/2Km6J6X
[12] C. Mao; C. Liang; W. Luo; J. Bao; J. Shen; X. Houa; Wenbo Zhao. Preparation of lotus-leaf-like polystyrene micro- and nanostructure films and its blood compatibility. J. Mater. Chem., 2009,19, 9025-9029
[13] B. K. Filshie: Insect Ultrastruct. 1982, 1, 281.
[14] J. Qian, D. Chi, R. Chai, J. For. Res. 2016, 27, 1391.
[15] R. J. Wootton. Syst. Entomol. 1979, 4, 81.
[16] S. R. Jongerius; D. Lentink. Exp. Mech. 2010, 50, 1323.
[17] P. Simonis; A. Bay; J.P. Vigneron. Physical Review E · Setembre 2009
[18] M. Sun; A. Liang; G. S. Watson; J. A. Watson; Y. Zheng; J. Ju; L. Jiang; PLoS One 2012, 7, e35056.
[19] D. Byun; J. Hong; J. H. Ko; Y. J. Lee; H. C. Park; B.-K. Byun; J. R. Lukes; J. Bionic Eng. 2009, 6, 63.
[20] O. Sato; S. Kubo; Z.-Z. Gu; Acc. Chem. Res. 2008, 42, 1.
[21] Y. Su; B. Ji; K. Zhang; H. Gao; Y. Huang; K. Hwang. Langmuir 2010, 26, 4984.
[22] J. Nickerl; R. Helbig; H.-J. Schulz; C. Werner; C. Neinhuis. Zoomorphology 2013, 132, 183.
[23] A. Cassie; S. Baxter; Trans. Faraday Soc. 1944, 40, 546.
[24] X. Gao; X. Yan; X. Yao; L. Xu; K. Zhang; J. Zhang; B. Yang; L. Jiang. Adv. Mater. 2007, 19, 2213.
[25] W. Bidlingmayer; J. Am. Mosq. Control Assoc. 1994, 10, 272.
[26] a) G. S. Watson; D. W. Green; B. W. Cribb; C. L. Brown; C. R. Meritt; M. J. Tobin; J. Vongsvivut; M. Sun; A.-P. Liang; J. A. Watson. ACS Appl. Mater. Interfaces 2017, 9, 24381; b) Lotus leaf, https://flic.kr/p/dX18gu (consulta: febrer 2018); c) Planthopper hindwing, https://flic.kr/p/Vr9Ney, (consulta: febrer 2018).
[27] C. Neinhuis; W. Barthlott. Ann. Bot. 1997, 79, 667.
[28] X. Gao; L. Jiang, Nature 2004, 432, 36.
