



L'increïble (i amagat) món dels insectes II
-
- Home
-
- 13 of 24

En la primera part de l'article "L'increïble (i amagat) món dels insectes", parlàvem de la presència d'aquests, comptats per milions, a les parts més insospitades del nostre planeta. Una de les claus de la seva supervivència i adaptabilitat es troba en el seu interior i es relaciona amb les micro i nanoestructures que trobem quan analitzem en profunditat els insectes. Aquestes estructures estan íntimament relacionades amb moltes propietats com l'adhesió, la locomoció o les seves propietats físiques i químiques (òptica, fotònica, hidrofobicitat, sensors i actuadors mecànics, acústics, òptics). Després de parlar en la primera part de l'adhesió, la superhidrofobicitat o de les seves ales, continuem ara complementant la informació amb noves propietats i imatges.
Sensors i producció de senyals mecànics
Els insectes realitzen multitud de tasques complexes en tota mena d'entorns, des de volar, apariar-se, buscar aliments, etc. Dur a terme aquestes tasques suposen la superació d'un gran hàndicap, el fet d'estar "atrapats" en un exoesquelet rígid similar al d'una armadura medieval. Aquest exoesquelet proporciona beneficis clau, però al seu torn planteja tota una sèrie de reptes, per detectar i interactuar de manera efectiva amb el món exterior. Per resoldre aquests problemes, els insectes han desenvolupat òrgans mecanosensorials que proporcionen retroalimentació espacial i sensible a l'entorn. Aquests òrgans són similars (i amb freqüència complementen) els apèndixs que els insectes usen per a la locomoció; es componen en gran part de cutícules disposades de tal manera que confereixen diverses propietats materials com força, elasticitat, durabilitat i ressonància vibratòria.

Detecció de senyals mecànics
La gran majoria d'aquestes eines mecanosensorials se sustenten en un mateix mecanisme de senyalització basat en gradients iònics, que també és emprat per les cèl·lules ciliades coclears (auditives) en organismes de vertebrats [29]. Aquest mecanisme es basa en dendrites. La dendrita d'una neurona mecanosensorial es troba dins d'una capa de cèl·lules epitelials altament resistent [30]. Quan la dendrita s'estimula mecànicament, els canals de potassi de transducció s'obren, despolaritzant ràpidament la neurona associada i alertant l'insecte de la presència d'un estímul, tot això en temps sorprenents que es mouen en l'escala dels microsegons (model de Hodgkin-Huxley) [31]. De manera anàloga a com els humans mapem els senyals de les retines humanes en una imatge visual completa, es creu que els insectes processen senyals mecanosensitius en el seu conjunt, la qual cosa probablement els permet interpretar corrents d'aire, moviments corporals, senyals de comunicació i característiques de la superfície com si es tractésd'un "mapa" [32]. Aquests mecanismes associen els aspectes estructurals dels mecanosensors dels insectes (per exemple, morfologia, elasticitat i posició anatòmica) amb les forces que transdueixen. Aquestes relacions estructura-força són un tema central en el desenvolupament de l'electrònica sensorial, que té aplicacions en pròtesis, robòtica i monitoratge biològic [33, 34]. Les morfologies d'òrgans més comunes en els insectes per transduir senyals són les estructures en forma de setes, també conegudes com a pèls tàctils. Aquests pèls consisteixen en aquests buits, cadascun dels quals està unit a una sola neurona sensorial, i actuen com a palanca que transmeten una força mecànica als corresponents canals de mecanotransducció [35]. Físicament, són gruixudes, acaben en punts afilats, i reben suport estructural addicional de cèl·lules especialitzades [36]. Cada cabell té selectivitat direccional que varia en funció de la seva morfologia, la tija, l'angle relatiu de la cutícula, i la localització i/o tipus de canal iònic que posseeixen. A la figura 1 observem una imatge de microscopi de rastreig (SEM) de l’espècie Osmia spinulosa i observem unes microestructures en forma de pèls rígids amb la teòrica tasca de transduir senyals.

Fig.1. Imatge de SEM realitzada als CCITUB de l'espècie Phyllomer cedida pel Museu de Ciències Naturals de Barcelona.
Detecció i producció de vibracions
La pèrdua d'audició afecta un de cada sis humans adults i és una de les afeccions cròniques més generalitzades en l'edat adulta [37]. És indubtable que existeix la necessitat de sistemes miniaturitzats i biològicament compatibles que puguin detectar de manera eficient i també produir sons en diferents rangs de freqüència. Aquests sistemes poden usar com a exemple els sistemes associats als insectes. Els insectes poden "escoltar" tota mena de sons a través d'una gran varietat d'estructures, molt poques de les quals s'assemblen a les orelles dels vertebrats. El so és una vibració que es propaga com una ona de pressió mecànica a través d'un mitjà de transmissió, com l'aire, l'aigua o un substrat sòlid. Com que la majoria dels insectes són animals terrestres, l'aire és el mitjà portador més prominent. És, no obstant això, un mitjà compressible de baixa densitat. Això significa que encara que les ones de so es propaguin una distància considerable a través de l'aire, la seva intensitat disminueix molt més de pressa que en els medis sòlids o líquids, una taxa exponencial ja descrita per la llei d'atenuació de Stokes [38]. Els òrgans que poden detectar sons en l'aire a gran distància (camp llunyà) són, per tant, molt més sensibles que aquells que només són receptius als sons o vibracions que es produeixen en el seu veïnatge immediat (camp pròxim).
Els detectors de camp pròxim són molt comuns en el món dels insectes, des dels òrgans de Johnston en la base de les antenes de mosquits fins als òrgans similars a antenes que es projecten cap enrere en les paneroles, fins i tot els pèls tàctils esmentats anteriorment [39]. Els òrgans del timpà en insectes (detectors de camp llunyà) poden detectar l'ona de pressió d'un camp de so des de més de 10 m de distància i freqüències que van des de 2 fins més de 100 000 Hz [40]. Aquests òrgans estan compostos per tres components: una membrana timpànica, un sac ple d'aire pressionat contra la membrana i un òrgan sensorial cordotonal [41–43].
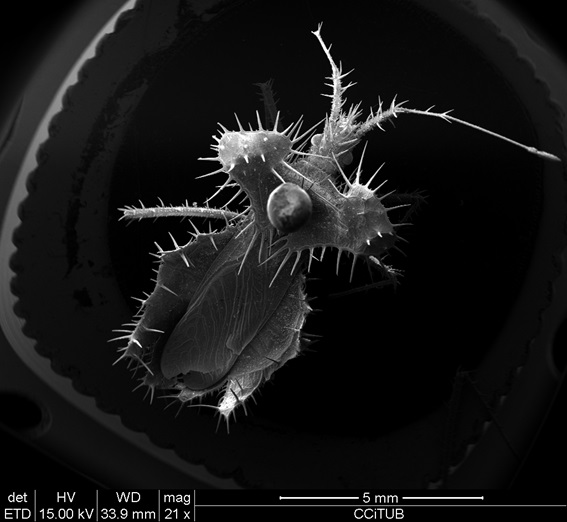
Figura 2. Imatge de SEM realitzada als CCITUB de l'espècie Phyllomer cedida pel Museu de Ciències Naturals de Barcelona.
Els insectes productors de llum: sensors i manipulació de llum
La majoria dels animals han utilitzat la llum com a portador principal d'informació per a la comunicació [44] des del sorgiment de la visió després de l'explosió en el Càmbric fa uns 500 milions d'anys [45, 46]. En particular, les intricades estructures òptiques lliuren senyals especialitzats que es processen en informació a través de sistemes visuals complexos, els ulls [47, 48]. L'exoesquelet de la cutícula de certs insectes conté nanoestructures ordenades, gairebé ordenades o desordenades que reflecteixen la llum en determinats rangs de longitud d'ona i poden produir colors cridaners, mentre que la cutícula en l'exterior d'altres insectes forma capes nanoestructurades que eviten la reflexió de la llum per complet, fent-les transparents. S'han desenvolupat mecanismes per manipular la llum que al mateix temps la detecten. La superfície d'alguns ulls d'insecte està modelada amb característiques en la nanoescala que promouen una transmissió eficient de la llum i també actuen com un element dissuasiu hidròfob per a la condensació que bloqueja la visió. Aquest conjunt d'eines de control de la llum és essencial per a la supervivència dels insectes i ha brindat inspiració per als sistemes d'enginyeria que aprofiten els fenòmens físics fonamentals per produir i detectar senyals visuals.
Mecanismes de producció de color
En general, existeixen dues classes principals de coloració animal: la coloració pigmentària a causa de l'absorció de llum selectiva per la longitud d'ona mitjançant tints químics, i la coloració estructural a causa de la interacció de la llum incident en nanoestructures ordenades, gairebé ordenades o desordenades que causen interferència [49-51] (figura 3). Tots dos mecanismes de coloració tenen propietats òptiques úniques que poden combinar de manera no trivial i modular tant propietats òptiques com aplicacions potencials, que van des de pantalles, fins a pintures brillants i duradores, fins a camuflatge adaptatiu i materials transparents [52-56].

Figura 3. L'ordre i el desordre en les estructures fotòniques poden portar a efectes òptics únics. A) Trastorn local en la cresta del reflector de la papallona, Morpho rhetenor, produeix un color blau estable. Adaptat amb permís. [57] Copyright 1999, la Royal Society. B) Estructura de Bouligand en escarabats joia, Chrysina gloriosa, dona com a resultat un senyal òptic polaritzat circularment. Adaptat [326] Copyright 2009, l'Associació Americana per a l'Avanç de la Ciència. C) Xarxa de quitina completament desordenada en escarabats blancs, l'estigma de Lepidiota resulta en una blancor brillant. Adaptat segons els termes de la llicència CC-BY [58] Copyright 2014, Matteo Burresi.
Transparència
Una altra forma usada pels insectes per evitar la detecció és emprant la transparència, proporcionant un camuflatge gairebé perfecte. Molts animals marins, com les meduses, són molt transparents [59]. No obstant això, aconseguir transparència és més fàcil amb il·luminació tènue o en aigua de mar tèrbola que en ambients ben il·luminats en l'aire, on s'observen fàcilment petits desajustaments de transparència. No obstant això, aquest efecte s'observa en les ales i els ulls de certs insectes, proporcionant una inspiració potencial per dissenyar materials transparents. La transparència és la propietat física que permet que la llum passi a través d'un material sense ser dispersada o absorbida. En termes més físics, la reflectivitat és mínima, mentre que la transmitància s'acosta a la unitat. El vidre és quasi transparent en el rang de longitud d'ona visible, però cada superfície de vidre és encara capaç de reflectir aproximadament el 4% de la llum incident a causa de la falta de coincidència de l'índex de refracció del vidre (RI ≈ 1,5) i l'aire (RI ≈ 1), la qual cosa resulta en una reflexió visible descrita per les equacions de Fresnel [59-61]. Per minimitzar aquest efecte, els materials en els sistemes naturals sovint s'estructuren de manera que l'índex de refracció canviï gradualment entre ells; això es coneix com a ajust d'impedància [60]. L'evolució ha seleccionat diversos trucs basats en nanoestructures per obtenir aquest efecte. Les ales transparents d'insectes són un exemple prominent d'igualació d'impedàncies. Si bé la majoria de les ales són pel·lícules primes de quitina [62], relativament poc estructurades, algunes libèl·lules, cigarrers i papallones han optimitzat la transparència de les seves ales emprant una nanoestructura local [337–341]. Un exemple el tenim en l'espècie Corythucha ciliata que adquireix transparència gràcies a les estructures que posseeix (figura 4).
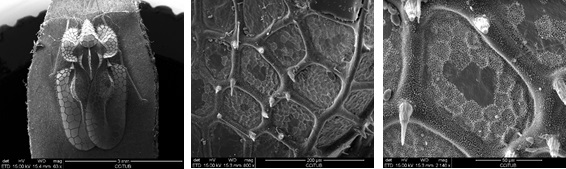
Figura 4 Imatges de SEM a diferents augments realitzades als CCITUB de l'espècie Corythucha ciliata cedides pel Museu de Ciències Naturals de Barcelona.
Els ulls dels insectes
A diferència dels humans, els insectes tenen ulls facetats i compostos que consisteixen en nombroses unitats anatòmicament idèntiques, com en els exemples de les espècies Tachinidae gonia i Phyllomer (figura 5) [68–72]. Es classifiquen segons el sistema òptic que empren per enfocar la llum incident de manera eficient en les parts sensibles a la llum de les seves cèl·lules fotoreceptores, ja sigui com a ulls de superposició òptica o d'aposició òptica. Els ulls d'aposició són el tipus principal d'ulls que es troben en els insectes. Aquí, cada faceta és un detector de llum separat. Consta d'una lent facetària que cobreix un ommatidi, que és un conjunt de cèl·lules fotoreceptores, cèl·lules pigmentàries i un con (cristal·lí) [73–75]. La llum incident s'enfoca a través de la lent en una llarga estructura cilíndrica similar a una guia d'ona anomenada rabdoma (fusionada). Els ulls de superposició òptica són emprats principalment per arnes nocturnes, i usen lents de múltiples facetes per enfocar la llum en fotoreceptores [73, 74, 76]. Els ulls d'aposició tenen una resolució espacial més alta que els ulls de superposició, mentre que els ulls de superposició tenen una major sensibilitat a la llum. Per tant, les arnes poden viure un estil de vida nocturn en contraposició amb les papallones, que estan actives durant el dia [68, 77]. El nombre de facetes per ull pot variar significativament, des de ≈100 en formigues fins a 30 000 en algunes libèl·lules. Un nombre baix de facetes fa que les formigues siguin gairebé incapaces de formar una imatge, mentre que les libèl·lules tenen una resolució superior.

Figura 5. Imatges de SEM realitzades als CCITUB de l'espècie a) Tachinidae gonia i b) Phyllomer cedides pel Museu de Ciències Naturals de Barcelona.
Sensors i regulació tèrmica
Les temperatures varien àmpliament en els diferents hàbitats on resideixen els insectes i les fluctuacions ocorren en un ampli rang d'escales de temps. Si bé la resposta als canvis de temperatura durant determinades èpoques (per exemple, les estacions) és important per a la hibernació i la supervivència de les espècies a llarg termini, els canvis a curt termini en la temperatura, que inclouen la nit i el dia i fins i tot el sol i l'ombra, són importants per a la supervivència d'alguns individus [78]. Els insectes empren solucions per als problemes de regulació tèrmica que abasten les disciplines científiques, des de controls biològics (per exemple, taxa metabòlica variable) fins a tècniques físiques (per exemple, coloració estructural i enfocaments químics (per exemple, síntesis de proteïnes anticongelants).
Sensors tèrmics
Alguns insectes han desenvolupat detectors de calor accionats mecànicament. Per exemple, els escarabats que busquen incendis forestals, Melanophila, volen als incendis forestals (i altres fonts de calor immensa) per dipositar els seus ous en arbres de coníferes recentment morts, ja que no poden superar la reacció de defensa resinosa natural d'arbres vius [79-80]. Els escarabats que busquen incendis forestals els detecten utilitzant els òrgans especialitzats en el pou de detecció d'infrarojos situats a banda i banda del tòrax, prop de les seves potes mitjanes. Cada òrgan sensorial consisteix en una estructura cuticular esfèrica ≈12–16 μm de diàmetre amb una cavitat central que està connectada al procés distal d'una cèl·lula nerviosa. Adjacent a cada òrgan sensorial, una glàndula neteja contínuament el sensor secretant regalims de cera [80]. La llum infraroja (IR) es detecta a través de l'absorció de llum en l'òrgan del pou, la qual cosa porta a una diferència de pressió detectada per un sensor de força [79-81]. En certa manera, aquests escarabats senten la llum IR. Els incendis forestals es cremen a temperatures entre 400 i 1200 °C, per la qual cosa emeten longituds d'ona IR en el rang de 2 a 4 μm [82]. Les neurones sensitives als infraroigs de Melanophila són capaces de detectar i respondre a longituds d'ona exclusivament en aquesta regió [83]. Aquestes troballes indiquen clarament que els escarabats Melanophila poden detectar un incendi de 10 hectàrees des d'una distància de 12 km a causa de la transparència atmosfèrica en aquestes longituds d'ona [79].

Control tèrmic
La distribució efectiva dels recursos alimentaris és de gran interès per a la creixent població mundial; més de 750 mil milions de dòlars d'aliments peribles es malgasten cada any, molts dels quals estan relacionats amb l'elevada temperatura dels aliments durant el transport i l'emmagatzematge [84]. Malgrat la seva petita mida, els insectes han trobat principis de disseny únics per regular la temperatura, els quals són biocompatibles. Existeixen dos tipus de mecanismes de termoregulació en els insectes; els que manipulen la producció de calor interna i els que manipulen l'intercanvi de calor externa. El vol està fonamentalment vinculat a la termoregulació, ja que el vol és una forma de locomoció energèticament costosa que generalment requereix una alta taxa metabòlica per subministrar suficient energia. Per volar, els músculs de vol d'un insecte han de ser capaços de produir una gran potència mecànica, que al seu torn produeix grans quantitats de calor. En condicions suaus, la calor generada per un insecte volador es dissipa sense causar cap mal. No obstant això, si l'insecte volador també està exposat a fonts externes de calor, com la llum solar o una temperatura ambient superior a la normal, ha de termoregular-se per mantenir una temperatura corporal no letal.
Un exemple de biomimetisme el tenim en l'empresa d'alimentació Unilever, la qual va presentar recentment una patent sobre la incorporació de proteïnes resistents a la congelació modificades genèticament per a la producció de gelats [85]. Amb l'addició de petites quantitats d'aquestes proteïnes, la companyia afirma que el gelat es pot congelar a temperatures inferiors a -40 °C sense cap risc de formació de cristalls de gel (que normalment provoquen cremades per congelació) quan es descongelen a la temperatura dels congeladors domèstics comuns (−4 a −16 °C). El gelat congelat trigaria molt més a fondre's si s'exposa a temperatures elevades d'enviament, la qual cosa redueix el risc de deterioració. Aquest concepte pot estendre's a altres substàncies peribles més nutritives.
Sensors químics i defensa
En major mesura que els vertebrats, els insectes naveguen pel seu món en gran part mitjançant la detecció i el desplegament d'estímuls químics. Els insectes usen l'olfacte per trobar el seu menjar i els seus companys [86], i produeixen molècules de senyalització volàtils anomenades feromones per transmetre una àmplia gamma de missatges als organismes que els envolten. No obstant això, els insectes han de superar una dificultat fonamental per enviar i rebre comunicacions químiques de manera efectiva: el món és gran i és petit. Els volums de les emissions de feromones o al·lomones d'un insecte solen ser d'un sol microlitre, com a màxim, per la qual cosa els compostos de senyalització volàtils es dilueixen extremadament amb l'evaporació. Com a resultat d'aquesta dilució, els insectes han desenvolupat sensors químics sofisticats que brinden a les neurones una exposició màxima al medi ambient al mateix temps que ofereixen protecció i selectivitat segons sigui necessari, així com diversos mètodes per dispersar els seus senyals químics de manera que garanteixi la proximitat als seus objectius. Els camps de detecció química i administració de fàrmacs també enfronten el problema de la dilució. En tots dos contextos, recentment s'han aplicat innovacions inspirades en insectes per superar-les. L'àmplia varietat de mecanismes dels insectes per a la detecció i dispersió química generalment involucra àrees de gran superfície i/o mitjans sofisticats per moure molècules a través de la pell o la cutícula, estratègies que també tenen utilitat en contextos dissenyats.

Conclusions
Hem compartit algunes de les característiques associades a les micro i nanoestructures que es troben en el fascinant i vast món dels insectes. Tenim l'esperança que els lectors puguin ara o en el futur identificar un problema que comparteixen amb un insecte que ha desenvolupat una solució eficient. L'article es basa en el review: It's Not a Bug, It's a Feature: Functional Materials in Insects de Schroeder et al., i representa una minúscula, diríem que nano, descripció d'alguns dels materials funcionals més atractius dels insectes, tenint en compte, tal com comentàvem abans, que existeixen més d'un milió d'espècies d'insectes conegudes i per tant, milers de possibilitats i característiques per a la seva completa descripció.
Fer una aproximació al món dels insectes és d'extremada importància, ja que tant l'abundància com la seva diversitat està en un alarmant declivi. La població general d'insectes ha disminuït en un 45% en els últims 40 anys, aquesta desacceleració s'està produint en el context d'una "sisena ona d'extinció" antropogènica del mateix ordre de magnitud que la suma de les cinc extincions en massa anteriors.
El valor total dels serveis prestats per insectes només als Estats Units s'ha estimat en 57 mil milions de dòlars anuals, la qual cosa, a més dels elements de línia obvis (com la pol·linització i l'alimentació d'animals més grans), també inclou beneficis menys visibles, com els 380 milions de dòlars que els escarabats piloters aporten als ranxers dels Estats Units cada any enterrant femta de bestiar. Donada la importància dels insectes en la cadena alimentària mundial i com a pol·linitzadors, les conseqüències d'aquestes disminucions poden ser nefastes, tant ecològicament com econòmicament.
Agraïments a la Dra. Aranzazu Villuendas, la Sra. Eva Prats i el Sr. J.M. Rebled dels Centres Científics i Tecnològics de la Universitat de Barcelona per l'obtenció de les imatges de microscòpia SEM i a Berta Caballero del Museu de Ciències Naturals de Barcelona per haver-nos deixat algunes de les mostres de l'estudi.
BIBLIOGRAFIA
[29] D. Corey; A. Hudspeth, Nature 1979, 281, 675.
[30] J. C. Tuthill; R. I. Wilson, Curr. Biol. 2016, 26, R1022.
[31] R. G. Walker; A. T. Willingham; C. S. Zuker, Science 2000, 287, 2229.
[32] G. A. Jacobs; J. P. Miller; Z. Aldworth; J. Exp. Biol. 2008, 211, 1819.
[33] T. Sekitani; T. Someya; Stretchable Electron (Ed: T. Someya). Wiley-VCH, Weinheim, Alemanya 2012, p. 271–285.
[34] L.-Q. Tao; K.-N. Zhang; H. Tian; Y. Liu; D.-Y. Wang; Y.-Q. Chen; Y. Yang; T.-L. Ren. ACS Nano 2017, 11, 8790.
[35] A. Chortos; G. Koleilat; R. Pfattner; D. Kong; L. Pei; R. Nur: T. Lei; H. Evan Wang; N. Liu; Y.-C. Lai; M.-G. Kim; J. W. Chung; S. Lee; Z. Bao. Adv. Mater. 2015, 28, 4441.
[36] U. Thurm. Cold Spring Harbor Symposia on Quantitative Biology, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, EUA 1965, p. 75–82.
[37] B. Shield. "Evaluation of the Social and Economic Costs of Hearing Impairment", A Report for Hear-It (Consulta: Febrer 2018).
[38] J. C. F. Chow. J. Acoust. Soc. Am. 1964, 36, 2395.
[39] O. Yamashita; K. Hasegawa. Comprehensive Insect Physiology, Biochemistry and Pharmacology, Vol. 1 (Eds: G. A. Kerkut, L. I. Gilbert), Pergamon Press, Oxford, Regne Unit 1985.
[40] M. B. Fenton; J. H. Fullard. J. Comp. Physiol., A 1979, 132, 77.
[41] L. A. Miller; J. Morphol. 1970, 131, 359.
[42] K. D. Roeder; J. Insect Physiol. 1972, 18, 1249.
[43] J. Schwabe. Beiträge Zur Morphologie Und Histologie Der Tympanalen Sinnesapparate Der Orthopteren (Ed: E. Nagele), E. Schweizerbartsche Verlagsbuchhandlung, Stuttgart, Germany1906.
[44] P. M. Lester. Visual Communication: Images with Messages,Wadsworth, Cengage Learning, Boston, MA, EUA 2013.
[45] A. R. Parker. J. Opt. A: Pure Appl. Opt. 2000, 2, R15.
[46] D.-E. Nilsson. Facets of Vision (Eds: D. G. Stavenga, R. C. Hardie), Springer, Berlin/Heidelberg, Germany 1989, p. 30–73.
[47] S. M. Doucet; M. G. Meadows. J. R. Soc., Interface 2009, 6 Suppl2, S115.
[48] I. C. Cuthill; W. L. Allen; K. Arbuckle; B. Caspers; G. Chaplin; M. E. Hauber; G. E. Hill; N. G. Jablonski; C. D. Jiggins; A. Kelber; J. Mappes; J. Marshall; R. Merrill; D. Osorio; R. Prum; N. W. Roberts; A. Roulin; H. M. Rowland; T. N. Sherratt; J. Skelhorn; M. P. Speed; M. Stevens; M. C. Stoddard; D. Stuart-Fox; L. Talas; E. Tibbetts; T. Caro. Science 2017, 357, eaan0221.
[49] S. Kinoshita. "Structural Colors in the Realm of Nature", World Scientific, Singapur 2008.
[50] G. I. Mark; Z. Vertesy; K. Kertesz; Z. Balint; L. P. Biro. Phys. Rev. E 2009, 80, 051903.
[51] L. P. Biro; J. P. Vigneron. Laser Photonics Rev. 2011, 5, 27.
[52] A. R. Parker; N. Martini. Opt. Laser Technol. 2006, 38, 315.
[53] P. Vukusic; J. R. Sambles. Nature 2003, 424, 852.
[54] A. L. Ingram; A. R. Parker. Philos. Trans. R. Soc., B 2008, 363, 2465.
[55] S. Kinoshita; S. Yoshioka; J. Miyazaki. Rep. Prog. Phys. 2008, 71, 076401. www.advancedsciencenews.com www.advmat.de 1705322 (44 of 48) c 2018 The Authors. Published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
[56] M. D. Shawkey; L. d’Alba. Philos. Trans. R. Soc., B 2017, 372, 20160536.
[57] P. Vukusic; J. R. Sambles; C. R. Lawrence; R. J. Wootton. Proc. R. Soc. Londres, Ser. B 1999, 266, 1403.
[58] M. Burresi; L. Cortese; L. Pattelli; M. Kolle; P. Vukusic; D. S. Wiersma; U. Steiner; S. Vignolini. Sci. Rep. 2014, 4, 6075.
[59] S. Johnsen. Sci. Am. 2000, 282, 80.
[60] E. Hecht. Optics, Addison-Wesley, Reading, MA, EUA 2001.
[61] Y. B. Band. Light and Matter: Electromagnetism, Optics, Spectroscopy and Lasers, Wiley, Chichester, Regne Unit 2006.
[62] E. Shevtsova; C. Hansson; D. H. Janzen; J. Kjaerandsen. Proc. Natl. Acad. Sci. EUA 2011, 108, 668.
[63] I. R. Hooper; P. Vukusic; R. J. Wootton. Opt. Express 2006, 14, 4891.
[64] a) Glass wing butterfly at Bristol Zoo, https://commons.wikimedia.org/wiki/Greta_morgane#/media/File:Glass_Wing_at_Bristol_Zoo.jpg , (consulta: febrer 2018); b) R. H. Siddique; G. Gomard; H. Holscher. Nat. Commun. 2015, 6, 6909.
{kind=link}
[65] G. Zhang; J. Zhang; G. Xie; Z. Liu; H. Shao. Small 2006, 2, 1440.
[66] L. Dellieu; M. Sarrazin; P. Simonis; O. Deparis; J. P. Vigneron. J. Appl. Phys. 2014, 116, 024701.
[67] G. S. Watson; J. A. Watson. Appl. Surf. Sci. 2004, 235, 139.
[68] a) Compound eyes of a robber fly, https://en.wikipedia.org/wiki/Holcocephala_fusca#/media/File:Robber_Fly_with_prey_(Holcocephala_fusca)_by_Thomas_Shahan.jpg, (consulta: febrer 2018); b) E. J. Warrant. Philos. Trans. R. Soc., B 2017, 372, 20160063.
_by_Thomas_Shahan.jpg){kind=link}
[69] P. Kunze. Comparative Physiology and Evolution of Vision in Invertebrates, Springer-Verlag, Berlin/Heidelberg, Alemanya 1979, p. 441–502.
[70] E. Warrant; D.-E. Nilsson. Invertebrate Vision, CambridgeUniversity Press, Cambridge, Regne Unit 2006.
[71] M. F. Land; R. D. Fernald. Annu. Rev. Neurosci. 1992, 15, 1.
[72] O. F. Lazareva; T. Shimizu; E. A. Wasserman. How Animals See the World: Comparative Behavior, Biology, and Evolution of Vision, Oxford University Press, Oxford, Regne Unit 2012.
[73] D. Grimaldi; M. S. Engel. Evolution of the Insects, Cambidge UniversityPress, Cambridge, Nova York, 2005.
[74] M. F. Land; D.-E. Nilsson. Animal Eyes, Oxford University Press, Oxford, Regne Unit 2012.
[75] A. W. Snyder. Comparative Physiology and Evolution of Visions in Invertebrates, Springer-Verlag. Berlín/Heidelberg, Alemanya 1979, p. 225–313.
[76] S. Exner. Die Physiologie Der Facettirten Augen von Krebsen UndInsecten: Eine Studie, Franz Deuticke, Leipzig, Alemanya 1891.
[77] A. Honkanen; E.-V. Immonen; I. Salmela; K. Heimonen; M. Weckstrom. Philos. Trans. R. Soc., B 2017, 372, 20160077.
[78] S. L. Chown; S. Nicolson. Insect Physiological Ecology: Mechanisms and Patterns, Oxford University Press, Oxford, Regne Unit 2004.
[79] a) Melanophila acuminata, https://de.wikipedia.org/wiki/Schwarzer_ Kiefernprachtk%C3%A4fer#/media/File:Melanophila_acuminata. jpg, (consulta: febrer 2018); b) H. Schmitz; H. Bleckmann. J. Comp. Physiol., A 1998, 182, 647.
[80] W. G. Evans. Ann. Entomol. Soc. Am. 1966, 59, 873.
[81] M. Muller; M. Olek; M. Giersig; H. Schmitz. J. Exp. Biol. 2008, 211, 2576.
[82] W. Gronenberg; H. Schmitz. Cell Tissue Res. 1999, 297, 311.
[83] W. G. Evans. Ecology 1966, 47, 1061.
[84] F. and A. O. of the U. Nations, Food Wastage Footprint: Impacts on Natural Resources: Summary Report, FAO, 2013.
[85] H. Y. Toschka; J. M. A. Verbakel; J. W. Almkerk. WO1994003617 A1, 1994.
